
Všetok obsah iLive je lekársky kontrolovaný alebo kontrolovaný, aby sa zabezpečila čo najväčšia presnosť faktov.
Máme prísne smernice týkajúce sa získavania zdrojov a len odkaz na seriózne mediálne stránky, akademické výskumné inštitúcie a vždy, keď je to možné, na lekársky partnerské štúdie. Všimnite si, že čísla v zátvorkách ([1], [2] atď.) Sú odkazmi na kliknutia na tieto štúdie.
Ak máte pocit, že niektorý z našich obsahov je nepresný, neaktuálny alebo inak sporný, vyberte ho a stlačte kláves Ctrl + Enter.
Hypotalamus
Lekársky expert článku

Posledná kontrola: 07.07.2025
Hypotalamus tvorí dolné časti diencefalonu a podieľa sa na tvorbe dna tretej komory. Hypotalamus zahŕňa optickú chiasmu, optický trakt, sivý tuberkul s lievikom a mamilárne telieska.
Optická chiasma (chiasma opticum) je priečny hrebeň tvorený vláknami zrakových nervov (II. pár hlavových nervov), čiastočne prechádzajúcimi na opačnú stranu (tvoriacimi kríženie). Tento hrebeň pokračuje laterálne a dozadu na každej strane do optického traktu (tratus opticus). Optický trakt sa nachádza mediálne a dozadu od prednej perforovanej substancie, ohýba sa z laterálnej strany okolo mozgovej stopky a končí dvoma koreňmi v subkortikálnych zrakových centrách. Väčší laterálny koreň (radix lateralis) sa približuje k laterálnemu genikulárnemu teliesku a tenší mediálny koreň (radix medialis) smeruje k hornému kolikulu strechy stredného mozgu.
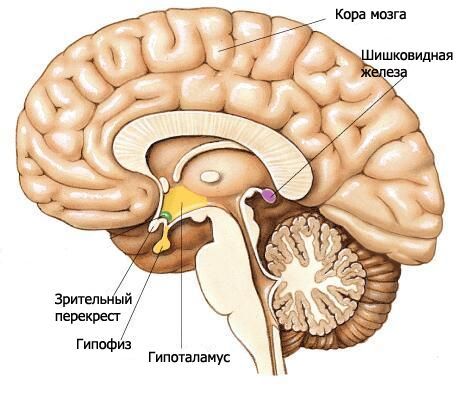
Terminálna platnička, ktorá patrí telencefalonu, prilieha k prednému povrchu optickej chiasmy a s ňou sa srastá. Uzatvára prednú časť pozdĺžnej štrbiny mozgu a pozostáva z tenkej vrstvy sivej hmoty, ktorá v laterálnych častiach platničky pokračuje do substancie čelových lalokov hemisfér.
Za optickou chiasmou sa nachádza sivý tuberkul (tuber cinereum), za ktorým ležia mamilárne telieska a po stranách sú optické dráhy. Pod ním sivý tuberkul prechádza do lievika (infundibulum), ktorý sa spája s hypofýzou. Steny sivého tuberkulu sú tvorené tenkou platničkou sivej hmoty obsahujúcou sivo-tuberálne jadrá (nuclei tuberales). Zo strany dutiny tretej komory vyčnieva do oblasti sivého tuberkulu a ďalej do lievika zužujúca sa priehlbina lievika.
Mamilárne telieska (corpora mamillaria) sa nachádzajú medzi sivým tuberkulom vpredu a zadnou perforovanou hmotou vzadu. Vyzerajú ako dva malé, guľovité biele útvary s priemerom približne 0,5 cm. Biela hmota sa nachádza iba mimo mamilárneho tela. Vo vnútri sa nachádza sivá hmota, v ktorej sa rozlišujú mediálne a laterálne jadrá mamilárneho tela (nuclei corporis mamillaris mediales et laterales). Stĺpce fornixu končia v mamilárnych telieskach.
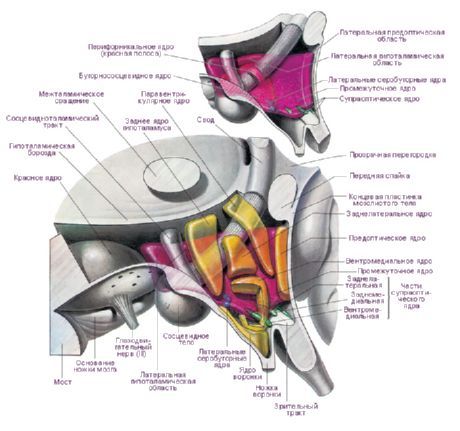
V hypotalame sa nachádzajú tri hlavné hypotalamické oblasti - zhluky skupín nervových buniek rôznych tvarov a veľkostí: predná (regio hypothalamica anterior), stredná (regio hypothalamica intermedia) a zadná (regio hypothalamica posterior). Zhluky nervových buniek v týchto oblastiach tvoria viac ako 30 jadier hypotalamu.
Nervové bunky hypotalamických jadier majú schopnosť produkovať sekrét (neurosekréciu), ktorý môže byť transportovaný do hypofýzy prostredníctvom výbežkov týchto buniek. Takéto jadrá sa nazývajú neurosekrečné jadrá hypotalamu. V prednej oblasti hypotalamu sa nachádza supraoptické (supraoptické) jadro (nucleus supraopticus) a paraventrikulárne jadrá (nuclei paraventriculares). Výbežky buniek týchto jadier tvoria hypotalamo-hypofyzárny zväzok, ktorý končí v zadnom laloku hypofýzy. Spomedzi skupiny jadier zadnej oblasti hypotalamu sú najväčšie mediálne a laterálne jadrá mamilárneho tela (nuclei corporis mamillaris mediales et laterales) a zadné hypotalamické jadro (nucleus hypothalamicus posterior). Skupina jadier strednej hypotalamickej oblasti zahŕňa inferomediálne a superomediálne hypotalamické jadrá (nuclei hypothalamic ventromediales et dorsomediales), dorzálne hypotalamické jadro (nucleus hypothalamicus dorsalis), jadro infundibulárneho svalu (nucleus infundibularis), sivohľuznaté jadrá (nuclei tuberales) atď.
Jadrá hypotalamu sú prepojené pomerne zložitým systémom aferentných a eferentných dráh. Hypotalamus má preto regulačný účinok na mnohé vegetatívne funkcie tela. Neurosekrécia jadier hypotalamu môže ovplyvniť funkcie žľazových buniek hypofýzy, zvýšiť alebo inhibovať sekréciu mnohých hormónov, ktoré následne regulujú aktivitu iných endokrinných žliaz.
Prítomnosť nervových a humorálnych spojení medzi hypotalamickými jadrami a hypofýzou umožnila ich spojenie do hypotalamo-hypofyzárneho systému.
Fylogenetické štúdie ukázali, že hypotalamus existuje u všetkých chordátov, je dobre vyvinutý u obojživelníkov a ešte viac u plazov a rýb. Vtáky majú jasne vyjadrenú diferenciáciu jadier. U cicavcov je sivá hmota vysoko vyvinutá, ktorej bunky sa diferencujú na jadrá a polia. Ľudský hypotalamus sa významne nelíši od hypotalamu vyšších cicavcov.
Existuje veľké množstvo klasifikácií hypotalamických jadier. E. Gruntel identifikoval 15 párov jadier, W. Le Gros Clark - 16, H. Kuhlenbek - 29. Najpoužívanejšou klasifikáciou je klasifikácia W. Le Gros Clarka. IN Bogolepova (1968) na základe vyššie uvedených klasifikácií a s prihliadnutím na údaje o ontogenéze navrhuje rozdeliť hypotalamické jadrá do štyroch častí:
- predná alebo rostrálna časť (kombinujúca preoptickú oblasť a prednú skupinu - W. Le Gros Clark) - preoptická mediálna a laterálna oblasť, suprachiasmatické jadro, supraoptické jadro, paraventrikulárne jadro, predná hypotalamická oblasť;
- stredná mediálna časť - ventromediálne jadro, dorzomediálne jadro, infundibulárne jadro, zadná hypotalamická oblasť;
- stredná laterálna časť - laterálna hypotalamická oblasť, laterálne hypotalamické jadro, tuberolaterálne jadro, tuberomamilárne jadro, perifornikálne jadro;
- zadná alebo mamilárna časť - mediálne mamilárne jadro, laterálne mamilárne jadro.
Anatomické súvislosti hypotalamu tiež objasňujú jeho (funkčný) význam. Medzi najdôležitejšie aferentné dráhy možno rozlíšiť:
- mediálny zväzok predného mozgu, ktorého laterálna časť spája hypotalamus s čuchovou bulbou a tuberkulom, periamygdaloidnou oblasťou a hipokampom a mediálna časť so septom, diagonálnou oblasťou a nucleus caudatus;
- terminálny pás, ktorý vedie od mandlí k predným častiam hypotalamu;
- vlákna prechádzajúce cez fornix z hipokampu do tela mamilára;
- talamo-, strio- a pallidohypotalamické spojenia;
- z mozgového kmeňa - centrálny tegmentálny trakt;
- z mozgovej kôry (orbitálnej, temporálnej, parietálnej).
Hlavnými zdrojmi aferentácie sú teda limbické formácie predného mozgu a retikulárna formácia mozgového kmeňa.
Eferentné systémy hypotalamu možno tiež rozdeliť do troch smerov:
- zostupné systémy k retikulárnej formácii a mieche - periventrikulárny systém vlákien končiacich v strednom mozgu (pozdĺžny zadný zväzok), v autonómnych centrách kaudálneho kmeňa a miechy a mamilárno-tegmentálny zväzok, ktorý prechádza od mamilárnych teliesok k retikulárnej formácii stredného mozgu;
- dráhy do talamu z mamilárnych teliesok (mammathalamický zväzok), ktoré sú súčasťou uzavretého funkčného limbického systému;
- dráhy do hypofýzy - hypotalamo-hypofyzárna dráha z paraventrikulárnych (10-20% vlákien) a supraoptických (80-90%) jadier do zadných a čiastočne stredných lalokov hypofýzy, tuberohypofyzárna dráha z ventromediálnych a infundibulárnych jadier do adenohypofýzy.
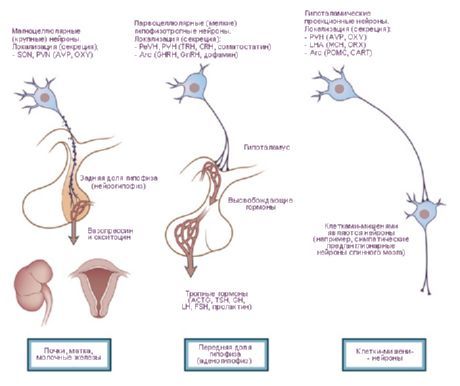
Práce J. Ransona (1935) a W. Hessa (1930, 1954, 1968) prezentovali údaje o rozširovaní a zúžení zrenice, zvýšení a znížení arteriálneho tlaku, zrýchlení a spomalení pulzu pri stimulácii hypotalamu. Na základe týchto štúdií boli identifikované zóny, ktoré vykazujú sympatické (zadná časť hypotalamu) a parasympatické (predná časť) účinky, a samotný hypotalamus bol považovaný za centrum integrujúce aktivitu viscerálneho systému inervujúceho orgány a tkanivá. S rozvojom týchto štúdií sa však objavil aj veľký počet somatických účinkov, najmä pri voľnom správaní zvierat [Gellhorn E., 1948]. O. G. Baklavadžan (1969) pri stimulácii rôznych častí hypotalamu pozoroval v niektorých prípadoch aktivačnú reakciu v mozgovej kôre, facilitáciu monosynaptických potenciálov miechy, zvýšenie arteriálneho tlaku a v iných opačný účinok. V tomto prípade mali vegetatívne reakcie najvyšší prah. O. Sager (1962) objavil inhibíciu y-systému a synchronizáciu EEG počas diatermie hypotalamu a opačný účinok pri nadmernom zahrievaní. Formuje sa predstava hypotalamu ako časti mozgu, ktorá vykonáva interakciu medzi regulačnými mechanizmami, integráciu somatickej a vegetatívnej aktivity. Z tohto hľadiska je správnejšie rozdeliť hypotalamus nie na sympatickú a parasympatickú časť, ale rozlišovať v ňom dynamogénne (ergotropné a trofotropné) zóny. Táto klasifikácia má funkčný, biologický charakter a odráža účasť hypotalamu na realizácii holistických behaviorálnych aktov. Je zrejmé, že na udržiavaní homeostázy sa podieľa nielen vegetatívny, ale aj somatický systém. Ergotropné a trofotropné zóny sa nachádzajú vo všetkých častiach hypotalamu a v niektorých oblastiach sa prekrývajú. Zároveň je možné identifikovať zóny ich „koncentrácie“. V predných častiach (preoptická zóna) sú teda trofotropné aparáty jasnejšie zastúpené a v zadných častiach (mamilárne telieska) ergotropné. Analýza hlavných aferentných a eferentných spojení hypotalamu s limbickým a retikulárnym systémom objasňuje jeho úlohu v organizácii integračných foriem správania. Hypotalamus zaujíma v tomto systéme osobitné - centrálne - postavenie, a to ako vďaka svojej topografickej polohe v strede týchto útvarov, tak aj v dôsledku fyziologických vlastností. Tieto sú určené úlohou hypotalamu ako špecificky skonštruovanej časti mozgu, obzvlášť citlivej na zmeny vo vnútornom prostredí tela, reagujúcej na najmenšie výkyvy humorálnych ukazovateľov a formujúcej účelné behaviorálne akty v reakcii na tieto zmeny.Špecifická úloha hypotalamu je predurčená jeho anatomickou a funkčnou blízkosťou k hypofýze. Jadrá hypotalamu sa delia na špecifické a nešpecifické. Prvá skupina zahŕňa útvary vyčnievajúce do hypofýzy, zvyšok zahŕňa iné jadrá, ktorých účinky stimulácie sa môžu líšiť v závislosti od sily nárazu. Špecifické jadrá hypotalamu majú zreteľný účinok a líšia sa od iných mozgových útvarov svojou schopnosťou neurokrinie. Patria sem supraoptické, paraventrikulárne a parvocelulárne jadrá sivého tuberkulu. Bolo zistené, že antidiuretický hormón (ADH) sa tvorí v supraoptických a paraventrikulárnych jadrách, ktoré zostupujú pozdĺž axónov hypotalamo-hypofyzárneho traktu do zadného laloku hypofýzy. Neskôr sa ukázalo, že v neurónoch hypotalamu sa tvoria uvoľňujúce faktory, ktoré po vstupe do adenohypofýzy regulujú sekréciu troch hormónov: adrenokortikotropného (ACTH), luteinizačného (LH), folikuly stimulujúceho (FSH) a tyreostimulujúceho (TSH). Zóny tvorby implementačných faktorov pre ACTH a TSH sú jadrá prednej časti mediánovej eminencie a preoptická oblasť a pre GTG - zadné časti sivého tuberkulu. Bolo zistené, že hypotalamo-hypofyzárne zväzky u ľudí obsahujú približne 1 milión nervových vlákien.
Nepochybne sa na neuroendokrinnej regulácii podieľajú aj iné časti mozgu (mediálno-bazálne štruktúry temporálnej oblasti, retikulárna formácia mozgového kmeňa). Najšpecifickejším aparátom je však hypotalamus, ktorý zahŕňa endokrinné žľazy v systéme integrálnych reakcií tela, najmä reakcií stresového charakteru. Trofo- a ergotropné systémy majú k dispozícii nielen periférny sympatický a parasympatický systém na zabezpečenie aktivity, ale aj špecifické neurohormonálne aparáty. Hypotalamo-hypofyzárny systém, fungujúci na princípe spätnej väzby, je do značnej miery samoregulačný. Aktivitu tvorby implementačných faktorov určuje aj hladina hormónov v periférnej krvi.
Hypotalamus je teda dôležitou súčasťou limbického a retikulárneho systému mozgu, ale keďže je súčasťou týchto systémov, zachováva si svoje špecifické „vstupy“ vo forme špeciálnej citlivosti na zmeny vo vnútornom prostredí, ako aj špecifické „výstupy“ cez hypotalamo-hypofyzárny systém, paraventrikulárne spojenia s vegetatívnymi formáciami ležiacimi pod nimi, ako aj cez talamus a retikulárnu formáciu mozgového kmeňa do kôry a miechy.
 [
[ Čo je potrebné preskúmať?