
Všetok obsah iLive je lekársky kontrolovaný alebo kontrolovaný, aby sa zabezpečila čo najväčšia presnosť faktov.
Máme prísne smernice týkajúce sa získavania zdrojov a len odkaz na seriózne mediálne stránky, akademické výskumné inštitúcie a vždy, keď je to možné, na lekársky partnerské štúdie. Všimnite si, že čísla v zátvorkách ([1], [2] atď.) Sú odkazmi na kliknutia na tieto štúdie.
Ak máte pocit, že niektorý z našich obsahov je nepresný, neaktuálny alebo inak sporný, vyberte ho a stlačte kláves Ctrl + Enter.
Vaječník
Lekársky expert článku

Posledná kontrola: 04.07.2025

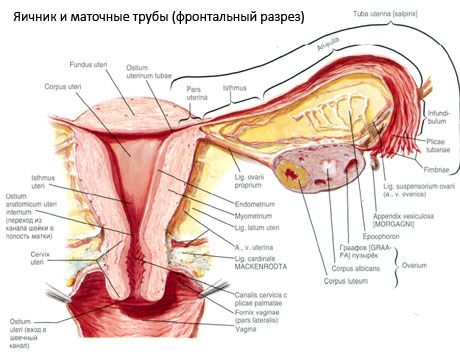
Vaječník (ovarium; grécky oophoron) je párový orgán, ženská reprodukčná žľaza, ktorá sa nachádza v panvovej dutine za širokým väzom maternice. Vo vaječníkoch sa vyvíjajú a dozrievajú ženské reprodukčné bunky (vajíčka) a tvoria sa ženské pohlavné hormóny, ktoré vstupujú do krvi a lymfy. Vaječník má vajcovitý tvar, v predozadnom smere je mierne sploštený. Farba vaječníka je ružovkastá. Na povrchu vaječníka ženy, ktorá rodila, sú viditeľné priehlbiny a jazvy - stopy ovulácie a premeny žltého telieska. Vaječník váži 5-8 g. Rozmery vaječníka sú: dĺžka 2,5-5,5 cm, šírka 1,5-3,0 cm, hrúbka - do 2 cm. Vaječník má dva voľné povrchy: mediálny povrch (facies medialis), otočený do panvovej dutiny, čiastočne pokrytý vajíčkovodom, a bočný povrch (facies lateralis), priliehajúci k bočnej stene panvy, k mierne vyjadrenému priehlbeniu - vaječníkovej jamke. Táto jamka sa nachádza v uhle medzi vonkajšími iliakálnymi cievami pokrytými peritoneom na vrchu a maternicovými a obturátorovými tepnami na spodnej strane. Za vaječníkom prechádza ureter príslušnej strany retroperitoneálne zhora nadol.
Povrchy vaječníkov prechádzajú do konvexného voľného (zadného) okraja (margo liber), spredu do mezenterického okraja (margo mesovaricus), ktorý je pripevnený krátkym záhybom pobrušnice (mezentéria vaječníka) k zadnému cípu širokého väzu maternice. Na tomto prednom okraji orgánu sa nachádza ryhovaná priehlbina - hilum vaječníkov (hilum ovarii), cez ktorý do vaječníka vstupujú tepny a nervy a vystupujú žily a lymfatické cievy. Vaječník má tiež dva konce: zaoblený horný tubulárny koniec (extremitas tubaria), smerujúci k vajíčkovodu, a dolný maternicový koniec (extremitas utenna), spojený s maternicou vaječníkovým väzom (lig. ovarii proprium). Tento väz, v tvare okrúhleho povrazca hrubého asi 6 mm, vedie od maternicového konca vaječníka k bočnému uhlu maternice, ktorý sa nachádza medzi dvoma listami širokého väzu maternice. Väzobný aparát vaječníka zahŕňa aj väzivo zavesujúce vaječník (lig.suspensorium ovarii), čo je záhyb pobrušnice, ktorý vedie od steny malej panvy k vaječníku a vo vnútri obsahuje cievy vaječníkov a zväzky vláknitých vlákien. Vaječník je upevnený krátkym mezentériom (mesovarium), čo je duplikát pobrušnice, ktorý vedie od zadného cípu širokého väzu maternice k mezenterickému okraju vaječníka. Samotné vaječníky nie sú pokryté pobrušnicou. Najväčší vaječníkový okraj vajcovodu je pripojený k tubulárnemu koncu vaječníka. Topografia vaječníka závisí od polohy maternice, jej veľkosti (počas tehotenstva). Vaječníky sú veľmi pohyblivé orgány dutiny malej panvy.
 [
[ Cievy a nervy vaječníkov
Krvné zásobenie vaječníkov zabezpečujú aa. et vv. ovaricae et uterinae. Obe ovariálne tepny (aa. ovaricae dextra et sinistra) vychádzajú z predného povrchu aorty tesne pod renálnymi tepnami; pravá zvyčajne vychádza z aorty a ľavá z renálnej tepny. Každá ovariálna tepna smeruje nadol a laterálne pozdĺž predného povrchu veľkého bedrového svalu (psoas major), križuje močovod spredu (od neho odvádza vetvy), vonkajšie iliakálne cievy, hraničnú čiaru a vstupuje do panvovej dutiny, kde sa nachádza v závesnom väze vaječníka. V mediálnom smere prechádza ovariálna tepna medzi listami širokého väzu maternice pod vajcovodom, od neho odvádza vetvy, a potom do mezentéria vaječníka; vstupuje do chilúma vaječníka.
Vetvy ovariálnej tepny sa široko anastomujú s ovariálnymi vetvami maternicovej tepny. Žilový odtok z vaječníkov sa uskutočňuje predovšetkým do ovariálneho venózneho plexu, ktorý sa nachádza v oblasti ovariálneho hilu. Odtiaľto odtok krvi prebieha dvoma smermi: cez maternicové a ovariálne žily. Pravá ovariálna žila má chlopne a vlieva sa do dolnej dutej žily. Ľavá ovariálna žila sa vlieva do ľavej renálnej žily, hoci nemá žiadne chlopne.
Lymfatická drenáž z vaječníkov prebieha cez lymfatické cievy, najmä hojne v oblasti orgánových brán, kde sa rozlišuje subovariálny lymfatický plexus. Lymfa sa potom odvádza pozdĺž ovariálnych lymfatických ciev do paraaortálnych lymfatických uzlín.
Inervácia vaječníkov
Sympatický - zabezpečený postgangliovými vláknami z celiakie (solárneho), horného mezenterického a hypogastrického plexu; parasympatický - viscerálnymi sakrálnymi nervami.
Štruktúra vaječníkov
Povrch vaječníka je pokrytý jednovrstvovým zárodočným epitelom. Pod ním sa nachádza hustý proteínový obal spojivového tkaniva (tunica albuginea). Spojivové tkanivo vaječníka tvorí jeho strómu (stroma ovarii), bohatú na elastické vlákna. Hmota vaječníka, jeho parenchým, sa delí na vonkajšiu a vnútornú vrstvu. Vnútorná vrstva, nachádzajúca sa v strede vaječníka, bližšie k jeho bráne, sa nazýva dreň (medulla ovarii). V tejto vrstve, v riedkom spojivovom tkanive, sa nachádzajú početné krvné a lymfatické cievy a nervy. Vonkajšia vrstva vaječníka - kôra (cortex ovarii) je hustejšia. Obsahuje veľa spojivového tkaniva, v ktorom sa nachádzajú dozrievajúce primárne ovariálne folikuly (folliculi ovarici primarii), sekundárne (vezikulárne) folikuly (folliculi ovarici secundarii, s.vesiculosi), ako aj zrelé folikuly, Graafove folikuly (folliculi ovarici maturis), ako aj žlté a atretické telieska.
Každý folikul obsahuje samičie reprodukčné vajíčko alebo oocyt (ovocytus). Vajíčko má priemer až 150 µm, je okrúhle, obsahuje jadro, veľké množstvo cytoplazmy, ktorá okrem bunkových organel obsahuje aj bielkovinovo-lipidové inklúzie (žĺtok), glykogén, potrebné na výživu vajíčka. Vajíčko zvyčajne spotrebuje svoju zásobu živín do 12 – 24 hodín po ovulácii. Ak k oplodneniu nedôjde, vajíčko odumiera.
Ľudské vajíčko má dve membrány. Vo vnútri sa nachádza cytoléma, čo je cytoplazmatická membrána vajíčka. Mimo cytolémy sa nachádza vrstva takzvaných folikulárnych buniek, ktoré chránia vajíčko a vykonávajú funkciu tvorby hormónov – vylučujú estrogény.
Fyziologickú polohu maternice, vajíčkovodov a vaječníkov zabezpečujú závesné, fixačné a podporné aparáty, ktoré spájajú pobrušnicu, väzy a panvové tkanivo. Závesný aparát je reprezentovaný párovými útvarmi, zahŕňa okrúhle a široké väzy maternice, vlastné väzy a závesné väzy vaječníkov. Široké väzy maternice, vlastné a závesné väzy vaječníkov držia maternicu v strednej polohe. Okrúhle väzy ťahajú fundus maternice dopredu a zabezpečujú jeho fyziologický sklon.
Fixačný (kotvový) aparát zaisťuje polohu maternice v strede malej panvy a prakticky znemožňuje jej posun do strán, tam a späť. Keďže však väzivový aparát odchádza od maternice v jej dolnej časti, sú možné naklonenia maternice v rôznych smeroch. Fixačný aparát zahŕňa väzy umiestnené v voľnom tkanive panvy a siahajúce od dolnej časti maternice k bočným, predným a zadným stenám panvy: sakro-magické, kardinálne, uterovezikálne a vezikopubické väzy.
Okrem mezovaria sa rozlišujú aj tieto ovariálne väzy:
- závesný väz vaječníka, predtým označovaný ako infundibulopelvický väz. Je to záhyb pobrušnice s krvnými cievami (a. et v. ovarica) a lymfatickými cievami a nervami vaječníka, ktorý ním prechádza, natiahnutý medzi bočnou stenou panvy, bedrovou fasciou (v oblasti rozdelenia spoločnej bedrovej tepny na vonkajšiu a vnútornú) a horným (vajcovodovým) koncom vaječníka;
- Vlastný väz vaječníka prechádza medzi listami širokého maternicového väzu, bližšie k zadnému listu, a spája spodný koniec vaječníka s bočným okrajom maternice. Vlastný väz vaječníka je pripojený k maternici medzi začiatkom vajcovodu a okrúhlym väzom, za a nad ním. V hrúbke väzu prechádzajú rr. ovarii, čo sú koncové vetvy maternicovej tepny;
- appendikulárno-vaječníkový väz Väzbovitosť sa tiahne od mezenteria slepého čreva k pravému vaječníku alebo širokému väzu maternice vo forme záhybu pobrušnice. Väzbovitosť je nestála a pozoruje sa u 1/2 - 1/3 žien.
Podporný aparát je reprezentovaný svalmi a fasciou panvového dna, rozdelenými na dolnú, strednú a hornú (vnútornú) vrstvu.
Najsilnejšia je horná (vnútorná) svalová vrstva, ktorú predstavuje párový sval, ktorý zdvíha konečník. Pozostáva zo svalových zväzkov, ktoré sa vejárovito rozširujú od kostrče k panvovým kostiam v troch smeroch (pubococcygeus, iliococcygeus a ischiococcygeus). Táto vrstva svalov sa nazýva aj panvová bránica.
Stredná vrstva svalov sa nachádza medzi symfýzou, lonovou kosťou a sedacou kosťou. Stredná vrstva svalov - urogenitálna bránica - zaberá prednú polovicu panvového vývodu, cez ktorý prechádza močová trubica a vagína. V prednej časti medzi jej vrstvami sa nachádzajú svalové zväzky, ktoré tvoria vonkajší zvierač močovej trubice, v zadnej časti sú svalové zväzky, ktoré idú v priečnom smere - hlboký priečny sval hrádze.
Spodná (vonkajšia) vrstva svalov panvového dna pozostáva z povrchových svalov, ktorých tvar pripomína číslo 8. Patria sem bulbokavernózny sval, ischiokavernózny sval, vonkajší análny zvierač a povrchový priečny perineálny sval.
Ontogenéza vaječníkov
Proces rastu a atrézie folikulov začína v 20. týždni tehotenstva a do pôrodu zostáva vo vaječníkoch dievčaťa až 2 milióny oocytov. Do menarché sa ich počet znižuje na 300 tisíc. Počas celého obdobia reprodukčného života dosiahne zrelosť a ovuláciu maximálne 500 folikulov. Počiatočný rast folikulov nezávisí od stimulácie FSH, je obmedzený a k atrézii dochádza rýchlo. Predpokladá sa, že namiesto steroidných hormónov sú hlavným regulátorom rastu a atrézie primárnych folikulov lokálne autokrinné/parakrinné peptidy. Predpokladá sa, že proces rastu a atrézie folikulov nie je prerušený žiadnymi fyziologickými procesmi. Tento proces pokračuje vo všetkých vekových kategóriách vrátane vnútromaternicového obdobia a menopauzy a je prerušený tehotenstvom, ovuláciou a anovuláciou. Mechanizmus, ktorý spúšťa rast folikulov a ich počet v každom konkrétnom cykle, stále nie je jasný.
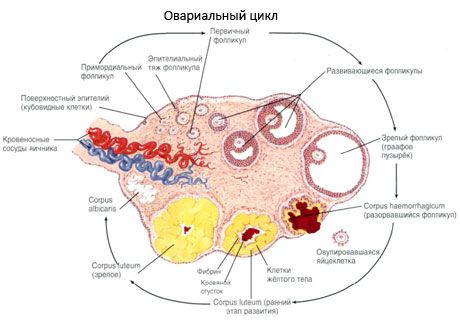
Počas svojho vývoja folikul prechádza niekoľkými vývojovými štádiami. Prvotné zárodočné bunky pochádzajú z endodermu žĺtkového vaku, alantoisu, a migrujú do genitálnej oblasti embrya v 5. – 6. týždni tehotenstva. V dôsledku rýchleho mitotického delenia, ktoré pokračuje od 6. – 8. týždňa do 16. – 20. týždňa tehotenstva, sa vo vaječníkoch embrya vytvorí až 6 – 7 miliónov oocytov, obklopených tenkou vrstvou granulóznych buniek.
Preantrálny folikul - oocyt je obklopený membránou (Zona pellucida). Granulosa bunky obklopujúce oocyt začínajú proliferovať, ich rast závisí od gonadotropínov a koreluje s hladinou estrogénov. Granulosa bunky sú cieľom FSH. V štádiu preantrálneho folikulu sú granulosa bunky schopné syntetizovať tri triedy steroidov: prevažne indukuje aktivitu aromatázy, hlavného enzýmu, ktorý premieňa androgény na estradiol. Predpokladá sa, že estradiol je schopný zvýšiť počet vlastných receptorov, čím vyvíja priamy mitogénny účinok na granulosa bunky nezávisle od FSH. Považuje sa za parakrinný faktor, ktorý zvyšuje účinky FSH vrátane aktivácie aromatizačných procesov.
Receptory FSH sa objavujú na membránach granulóznych buniek hneď po začatí rastu folikulu. Zníženie alebo zvýšenie FSH vedie k zmene počtu jeho receptorov. Tento účinok FSH je modulovaný rastovými faktormi. FSH pôsobí prostredníctvom G-proteínu, systému adenylátcyklázy, hoci steroidogenézu vo folikule reguluje hlavne FSH, do tohto procesu je zapojených mnoho faktorov: iónové kanály, tyrozínkinázové receptory, fosfolipázový systém sekundárnych poslov.
Úloha androgénov v skorom vývoji folikulov je zložitá. Granulózové bunky majú androgénne receptory. Tie nie sú len substrátom pre FSH-indukovanú aromatizáciu na estrogény, ale môžu tiež zosilniť aromatizáciu pri nízkych koncentráciách. Keď sa hladiny androgénov zvýšia, preantrálne granulózové bunky si prednostne nevyberajú aromatizačnú cestu na estrogény, ale jednoduchšiu androgénnu cestu cez 5α-reduktázu, aby sa premenili na androgén, ktorý sa nedá premeniť na estrogén, čím inhibujú aktivitu aromatázy. Tento proces tiež inhibuje tvorbu receptorov FSH a LH, čím sa zastavuje vývoj folikulov.
Proces aromatizácie, folikul s vysokou hladinou androgénov prechádza atréziou. Rast a vývoj folikulu závisí od jeho schopnosti premieňať androgény na estrogény.
V prítomnosti FSH budú dominantnou látkou folikulárnej tekutiny estrogény. V neprítomnosti FSH - androgény. LH normálne vo folikulárnej tekutine chýba až do polovice cyklu. Hneď ako sa hladina LH zvýši, mitotická aktivita granulóznych buniek sa zníži, objavia sa degeneratívne zmeny a hladina androgénov vo folikule sa zvýši. Hladina steroidov vo folikulárnej tekutine je vyššia ako v plazme a odráža funkčnú aktivitu ovariálnych buniek: granulóznych a thekálnych buniek. Ak je jediným cieľom FSH granulózne bunky, potom má LH mnoho cieľov - thekálne bunky, stromálne a luteínové bunky a granulózne bunky. Granulózne aj thekálne bunky majú schopnosť steroidogenézy, ale v granulóznych bunkách prevláda aromatázová aktivita.
V reakcii na LH produkujú thekálne bunky androgény, ktoré sú potom granulóznymi bunkami premieňané na estrogény prostredníctvom aromatizácie indukovanej FSH.
Ako folikul rastie, thekálne bunky začínajú exprimovať gény pre LH receptory, P450 sec a 3beta-hydroxysteroid dehydrogenázu, inzulínu podobný rastový faktor (IGF-1) synergicky s LH zvyšuje génovú expresiu, ale nestimuluje steroidogenézu.
Ovariálna steroidogenéza je vždy závislá od LH. Ako folikul rastie, theka bunky exprimujú enzým P450c17, ktorý tvorí androgény z cholesterolu. Granulózové bunky tento enzým nemajú a sú závislé od theka buniek, aby produkovali estrogény z androgénov. Na rozdiel od steroidogenézy je folikulogenéza závislá od FSH. Ako folikul rastie a hladiny estrogénu sa zvyšujú, aktivuje sa mechanizmus spätnej väzby - produkcia FSH je inhibovaná, čo následne vedie k zníženiu aktivity aromatázy folikulu a v konečnom dôsledku k atrézii folikulu prostredníctvom apoptózy (programovaná bunková smrť).
Mechanizmus spätnej väzby estrogénov a FSH inhibuje vývoj folikulov, ktoré začali rásť, ale nie dominantného folikulu. Dominantný folikul obsahuje viac receptorov FSH, ktoré podporujú proliferáciu granulóznych buniek a aromatizáciu androgénov na estrogény. Okrem toho parakrinná a autokrinná dráha pôsobia ako dôležitý koordinátor vývoja antrálneho folikulu.
Autokrinný/parakrinný regulátor pozostáva z peptidov (inhibín, aktivín, folistatín), ktoré sú syntetizované granulóznymi bunkami v reakcii na FSH a vstupujú do folikulárnej tekutiny. Inhibín znižuje sekréciu FSH; aktivín stimuluje uvoľňovanie FSH z hypofýzy a zvyšuje účinok FSH vo vaječníkoch; folistatín potláča aktivitu FSH, pravdepodobne väzbou na aktivín. Po ovulácii a vývoji žltého telieska je inhibín pod kontrolou LH.
Rast a diferenciáciu ovariálnych buniek ovplyvňujú inzulínu podobné rastové faktory (IGE). IGF-1 pôsobí na granulózne bunky, čo spôsobuje zvýšenie cyklického adenozínmonofosfátu (cAMP), progesterónu, oxytocínu, proteoglykánu a inhibínu.
IGF-1 pôsobí na theca bunky, čo spôsobuje zvýšenú produkciu androgénov. Theca bunky zase produkujú faktor nekrózy nádorov (TNF) a epidermálny rastový faktor (EGF), ktoré sú tiež regulované FSH.
EGF stimuluje proliferáciu granulóznych buniek. IGF-2 je hlavným rastovým faktorom vo folikulárnej tekutine a nachádzajú sa v nej aj IGF-1, TNF-a, TNF-3 a EGF.
Zdá sa, že narušenie parakrinnej a/alebo autokrinnej regulácie ovariálnej funkcie zohráva úlohu pri poruchách ovulácie a pri vývoji polycystických vaječníkov.
S rastom antrálneho folikulu sa zvyšuje obsah estrogénu vo folikulárnej tekutine. Na vrchole ich nárastu sa na granulóznych bunkách objavujú receptory pre LH, dochádza k luteinizácii granulóznych buniek a zvyšuje sa produkcia progesterónu. V preovulačnom období teda zvýšenie produkcie estrogénu spôsobuje objavenie sa LH receptorov, LH zase spôsobuje luteinizáciu granulóznych buniek a produkciu progesterónu. Zvýšenie progesterónu znižuje hladiny estrogénu, čo zrejme spôsobuje druhý vrchol FSH uprostred cyklu.
Predpokladá sa, že ovulácia nastáva 10 – 12 hodín po vrchole LH a 24 – 36 hodín po vrchole estradiolu. Predpokladá sa, že LH stimuluje redukciu delenia oocytov, luteinizáciu granulóznych buniek a syntézu progesterónu a prostaglandínov vo folikule.
Progesterón zvyšuje aktivitu proteolytických enzýmov, ktoré spolu s prostaglandínom podieľajú na prasknutí steny folikulu. Progesterónom indukovaný vrchol FSH umožňuje oocytu opustiť folikul premenou plazminogénu na proteolytický enzým plazmín a poskytuje dostatočný počet LH receptorov pre normálny vývoj luteálnej fázy.
Do 3 dní po ovulácii sa granulózne bunky zväčšujú a objavujú sa v nich charakteristické vakuoly naplnené pigmentom luteínom. Teka-luteálne bunky sa diferencujú z teky a strómy a stávajú sa súčasťou žltého telieska. Kapiláry prenikajúce do žltého telieska sa pod vplyvom faktorov angiogenézy vyvíjajú veľmi rýchlo a so zlepšenou vaskularizáciou sa zvyšuje produkcia progesterónu a estrogénov. Aktivita steroidogenézy a dĺžka života žltého telieska sú určené hladinou LH. Žlté teliesko nie je homogénny bunkový útvar. Okrem 2 typov luteálnych buniek obsahuje endotelové bunky, makrofágy, fibroblasty atď. Veľké luteálne bunky produkujú peptidy (relaxín, oxytocín) a sú aktívnejšie v steroidogenéze s väčšou aktivitou aromatázy a väčšou syntézou progesterónu ako malé bunky.
Vrchol progesterónu nastáva 8. deň po vrchole LH. Je potrebné poznamenať, že progesterón a estradiol sa v luteálnej fáze vylučujú epizodicky v korelácii s pulzatilnou produkciou LH. S tvorbou žltého telieska prechádza kontrola produkcie inhibínu z FSH na LH. Inhibín sa zvyšuje so zvýšením estradiolu pred vrcholom LH a naďalej sa zvyšuje aj po vrchole LH, hoci hladiny estrogénu klesajú. Hoci inhibín a estradiol sú vylučované granulóznymi bunkami, sú regulované rôznymi dráhami. Pokles inhibínu na konci luteálnej fázy prispieva k zvýšeniu FSH v nasledujúcom cykle.
Žlté teliesko sa veľmi rýchlo zmenšuje – 9. – 11. deň po ovulácii.
Mechanizmus degenerácie nie je jasný a nesúvisí s luteolytickou úlohou estrogénov ani s mechanizmom súvisiacim s receptormi, ako sa pozoruje v endometriu. Existuje aj iné vysvetlenie úlohy estrogénov produkovaných žltým telieskom (corpus luteum). Je známe, že estrogény sú potrebné na syntézu progesterónových receptorov v endometriu. Estrogény luteálnej fázy sú pravdepodobne nevyhnutné pre zmeny súvisiace s progesterónom v endometriu po ovulácii. Nedostatočný vývoj progesterónových receptorov v dôsledku nedostatočných hladín estrogénu je pravdepodobne ďalším mechanizmom neplodnosti a skorých potratov, čo je ďalšia forma deficitu luteálnej fázy. Predpokladá sa, že dĺžka života žltého telieska sa určuje v čase ovulácie. A určite dôjde k jeho regresii, ak nie je v dôsledku tehotenstva podporované ľudským choriovým gonadotropínom. Regresia žltého telieska teda vedie k zníženiu hladín estradiolu, progesterónu a inhibínu. Pokles inhibínu odstraňuje jeho supresívny účinok na FSH; Pokles estradiolu a progesterónu umožňuje veľmi rýchle obnovenie sekrécie GnRH a odstránenie mechanizmu spätnej väzby z hypofýzy. Pokles inhibínu a estradiolu spolu so zvýšením GnRH vedie k prevahe FSH nad LH. Zvýšenie FSH vedie k rastu folikulov s následným výberom dominantného folikulu a ak nedôjde k otehotneniu, začína sa nový cyklus. Steroidné hormóny zohrávajú vedúcu úlohu v reprodukčnej biológii a všeobecnej fyziológii. Určujú ľudský fenotyp, ovplyvňujú kardiovaskulárny systém, metabolizmus kostí, pokožku, celkovú pohodu a zohrávajú kľúčovú úlohu v tehotenstve. Pôsobenie steroidných hormónov odráža intracelulárne a genetické mechanizmy, ktoré sú potrebné na prenos extracelulárneho signálu do bunkového jadra, aby sa vyvolala fyziologická odpoveď.
Estrogény difúzne prenikajú bunkovou membránou a viažu sa na receptory nachádzajúce sa v bunkovom jadre. Komplex receptor-steroid sa potom viaže na DNA. V cieľových bunkách tieto interakcie vedú k génovej expresii, syntéze bielkovín a špecifickým funkciám buniek a tkanív.
Использованная литература