
Všetok obsah iLive je lekársky kontrolovaný alebo kontrolovaný, aby sa zabezpečila čo najväčšia presnosť faktov.
Máme prísne smernice týkajúce sa získavania zdrojov a len odkaz na seriózne mediálne stránky, akademické výskumné inštitúcie a vždy, keď je to možné, na lekársky partnerské štúdie. Všimnite si, že čísla v zátvorkách ([1], [2] atď.) Sú odkazmi na kliknutia na tieto štúdie.
Ak máte pocit, že niektorý z našich obsahov je nepresný, neaktuálny alebo inak sporný, vyberte ho a stlačte kláves Ctrl + Enter.
Príčiny a patogenéza záškrtu
Lekársky expert článku

Posledná kontrola: 04.07.2025
Záškrt je paradigmou toxigénnych infekčných chorôb. V roku 1883 Klebs preukázal, že pôvodcom záškrtu je Corynebacterium diphtheriae. O rok neskôr Loeffler zistil, že organizmus sa dá kultivovať iba z nosohltanovej dutiny a predpokladal, že poškodenie vnútorných orgánov je spôsobené rozpustným toxínom. V roku 1888 Roux a Yersin preukázali, že zvieratá, ktorým boli injekčne podané sterilné filtráty C. diphtheriae, vyvinuli orgánovú patológiu nerozoznateľnú od ľudskej záškrty; to preukázalo, že hlavným faktorom virulencie bol silný exotoxín.
Záškrt je najčastejšie infekcia horných dýchacích ciest, ktorá spôsobuje horúčku, bolesť hrdla a nevoľnosť. V mieste (miestach) infekcie sa často tvorí hrubá, sivozelená fibrínová membrána, pseudomembrána, v dôsledku kombinovaných účinkov bakteriálneho rastu, produkcie toxínov, nekrózy tkaniva a imunitnej odpovede hostiteľa. Zistenie, že systémové poškodenie orgánov je spôsobené pôsobením záškrtového toxínu, viedlo k vývoju účinnej terapie založenej na antitoxínoch na liečbu akútnej infekcie a vysoko účinnej toxoidovej vakcíny.
Hoci očkovanie toxoidmi urobilo zo záškrtu zriedkavé ochorenie v oblastiach, kde štandardy verejného zdravotníctva vyžadujú očkovanie, ohniská záškrtu sa stále vyskytujú u neočkovaných a imunokompromitovaných skupín. Naopak, v oblastiach, kde boli aktívne imunizačné programy pozastavené, sa vyskytli rozsiahle ohniská záškrtu dosahujúce epidemické rozmery.
Iné druhy korynebaktérií
Okrem C. diphtheriae, C. ulcerans a C. pseudotuberculosis môžu C. pseudodiphtheriticum a C. xerosis občas spôsobiť infekcie nosohltana a kože. Posledné dva kmene sú rozpoznateľné podľa ich schopnosti produkovať pyrazínamidázu. Vo veterinárnej medicíne sú C. renale a C. kutscheri dôležitými patogénmi, ktoré spôsobujú pyelonefritídu u hovädzieho dobytka a latentné infekcie u myší.
Príčiny
Pôvodca záškrtu, Corynebacterium diphtheriae, je tenká, mierne zakrivená tyčinka s kyjovitými zhrubnutiami na koncoch, nehybná; netvorí spóry, kapsuly ani bičíky a je grampozitívna.
Okrem toxínu produkujú difterické korynebaktérie počas svojich životných procesov neuraminidázu, hyaluronidázu, hemolyzín, nekrotizujúce a difúzne faktory, ktoré môžu spôsobiť nekrózu a skvapalnenie hlavnej látky spojivového tkaniva.
Na základe ich schopnosti tvoriť toxíny sa difterické korynebaktérie delia na toxigénne a netoxigénne.
Difterický toxín je silný bakteriálny exotoxín, ktorý určuje všeobecné aj lokálne klinické prejavy ochorenia. Toxigenicita je geneticky podmienená. Netoxigénne korynebaktérie diftérie ochorenie nespôsobujú.
Podľa kultúrnych a morfologických znakov sa všetky difterické korynebaktérie delia na 3 varianty: gravis, mitis, intermedius. Neexistuje priama závislosť závažnosti ochorenia od variantu difterických korynebaktérií. Každý variant obsahuje toxigénne aj netoxigénne kmene. Toxigénne korynebaktérie všetkých variantov produkujú identický toxín. [ 1 ]
Štruktúra, klasifikácia a typy antigénov
Corynebacterium diphtheriae je grampozitívna, nepohyblivá tyčinka v tvare kyja. Kmene rastúce v tkanive alebo staršie kultúry in vitro obsahujú v bunkových stenách jemné škvrny, ktoré umožňujú odfarbenie počas Gramovho farbenia a vedú k variabilnej Gramovej reakcii. Staršie kultúry často obsahujú metachromatické granule (polymetafosfát), ktoré sa farbia modrofialovo metylénovou modrou. Cukry bunkovej steny zahŕňajú arabinózu, galaktózu a manózu. Okrem toho sa môže izolovať toxický 6,6'-ester trehalózy, ktorý obsahuje korynemykolovú a korynemykolénovú kyselinu v ekvimolárnych koncentráciách. Rozoznávajú sa tri odlišné typy kultúr: mitis, intermedius a gravis.
Väčšina kmeňov potrebuje na rast kyselinu nikotínovú a pantoténovú; niektoré potrebujú aj tiamín, biotín alebo kyselinu pimelovú. Pre optimálnu produkciu difterického toxínu musí byť médium doplnené aminokyselinami a musí byť odložené.
Už v roku 1887 Loeffler opísal izoláciu avirulentných (netoxigénnych) kmeňov C. diphtheriae, ktoré boli nerozoznateľné od virulentných (toxigénnych) kmeňov izolovaných od pacientov u zdravých jedincov. V súčasnosti sa uznáva, že avirulentné kmene C. diphtheriae sa môžu po infekcii a lyzogenizácii jedným z mnohých odlišných korynebakteriofágov nesúcich štrukturálny gén pre difterický toxín, tox. Lyzogénna premena avirulentného na virulentný fenotyp sa môže vyskytnúť in situ aj in vitro. Štrukturálny gén pre difterický toxín nie je nevyhnutný ani pre korynebakteriofág, ani pre C. diphtheriae. Napriek tomuto pozorovaniu nebol pozorovaný genetický drift difterického toxínu.
 [
[ Patogenézy
Vstupnými bodmi infekcie sú sliznice orofaryngu, nosa, hrtana, menej často sliznica očí a genitálií, ako aj poškodená koža, rany alebo popáleniny, zapareniny, nezahojená pupočná ranka. V mieste vstupu sa difterická korynebaktéria množí a vylučuje exotoxín.
Exsudát bohatý na fibrinogén sa vylučuje a premieňa na fibrín pod vplyvom trombokinázy uvoľňovanej počas nekrózy epitelových buniek. Vytvára sa fibrínový film - charakteristický znak záškrtu.
Asymptomatické nazofaryngeálne nosičstvo je bežné v oblastiach endemických pre diftériu. U citlivých jedincov toxigénne kmene spôsobujú ochorenie replikáciou a vylučovaním difterického toxínu v nosohltane alebo kožných léziách. Difterická lézia je často pokrytá pseudomembránou zloženou z fibrínu, baktérií a zápalových buniek. Difterický toxín sa môže proteolyticky štiepiť na dva fragmenty: N-terminálny fragment A (katalytická doména) a fragment B (transmembránová a receptor-viažuca doména). Fragment A katalyzuje NAD+-dependentnú ADP-ribozyláciu elongačného faktora 2, čím inhibuje syntézu bielkovín v eukaryotických bunkách. Fragment B sa viaže na receptor na povrchu bunky a uľahčuje doručenie fragmentu A do cytozolu.
Ochranná imunita zahŕňa protilátkovú odpoveď na difterický toxín po klinickom ochorení alebo na difterický toxín (toxín inaktivovaný formaldehydom) po imunizácii.
Kolonizácia
O faktoroch, ktoré sprostredkúvajú kolonizáciu C. diphtheriae, sa vie len málo. Je však jasné, že k virulencii prispievajú aj iné faktory ako produkcia difterického toxínu. Epidemiologické štúdie ukázali, že daný lyzotyp môže v populácii pretrvávať dlhý čas. Neskôr môže byť nahradený iným lyzotypom. Vznik a následná dominancia nového lyzotypu v populácii pravdepodobne súvisí s jeho schopnosťou kolonizovať a efektívne konkurovať vo svojom segmente nazofaryngeálnej ekologickej niky. Corynebacterium diphtheriae môže produkovať neuraminidázu, ktorá štiepi kyselinu sialovú na povrchu buniek na jej zložky pyruvát a kyselinu N-acetylneuramínovú. Kordový faktor (6,6'-di-O-mykoloyl-α, α'-D-trehalóza) je povrchovou zložkou C. diphtheriae, ale jeho úloha pri kolonizácii ľudského hostiteľa nie je jasná.
Produkcia difterických toxínov
Štrukturálny gén difterického toxínu, tox, patrí do čeľade blízko príbuzných korynebakteriofágov, z ktorých β-fág je najlepšie preskúmaný. Regulácia expresie difterického toxínu je sprostredkovaná železom aktivovaným represorom DtxR, ktorý je kódovaný genómom C. diphtheriae. Expresia toxínu závisí od fyziologického stavu C. diphtheriae. Za podmienok, v ktorých sa železo stáva substrátom limitujúcim rýchlosť rastu, sa železo disociuje z DtxR, gén toxikodendrónu sa aktivuje a difterický toxín sa syntetizuje a vylučuje do kultivačného média maximálnou rýchlosťou.
Difterický toxín je nezvyčajne silný; pre vnímavé druhy (napr. ľudia, opice, králiky, morčatá) je smrteľná už dávka 100 až 150 ng/kg telesnej hmotnosti. Difterický toxín pozostáva z jedného polypeptidového reťazca s 535 aminokyselinami. Biochemická, genetická a röntgenová štrukturálna analýza ukazuje, že toxín pozostáva z troch štrukturálnych/funkčných domén:
- N-terminálna ADP-ribozyltransferáza (katalytická doména);
- oblasť, ktorá uľahčuje doručenie katalytickej domény cez bunkovú membránu (transmembránová doména);
- väzbová doména receptora eukaryotických buniek.
Po miernom trypsínovom rozštiepení a redukcii za denaturačných podmienok sa difterický toxín môže špecificky štiepiť v jeho proteázovo citlivej slučke na dva polypeptidové fragmenty (A a B). Fragment A je N-terminálna 21 kDa zložka toxínu a obsahuje katalytické miesto pre ADP-ribozyláciu elongačného faktora 2 (EF-2).
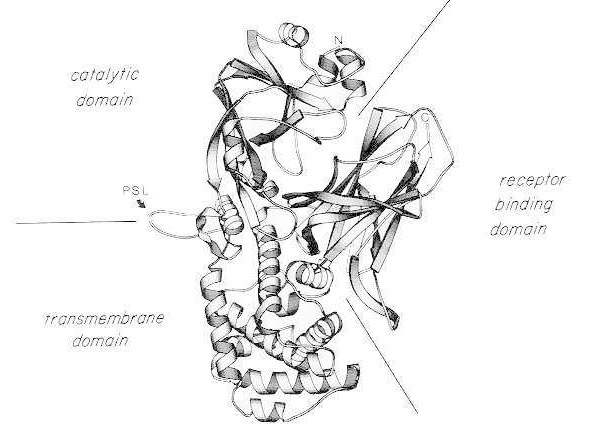
Páskový diagram röntgenovej kryštálovej štruktúry monomérneho natívneho difterického toxínu. (upravené z Bennett MJ, Choe S, Eisenberg D: Domain swapping: Entangling aliances between proteins. Proc Natl Acad Sci, USA, 91: 3127, 1994). Sú znázornené relatívne polohy katalytických, transmembránových a receptor-viažucich domén. Intaktný toxín môže byť štiepený trypsínovými proteázami na Arg190, Arg192 a/alebo Arg193, ktoré sa nachádzajú v proteázovo-senzitívnej slučke (PSL). Po redukcii disulfidového mostíka medzi Cys186 a Cys201 môže byť toxín rozdelený na fragmenty A a B. Sú znázornené amino-terminálne (N) a karboxy-terminálne (C) konco-intaktného toxínu. Páskový diagram bol vytvorený pomocou programu MOLESCRIPT.
C-terminálny fragment, fragment B, nesie transmembránovú a receptorovo-väzbovú doménu toxínu.
Otrava jednej eukaryotickej bunky difterickým toxínom zahŕňa najmenej štyri odlišné kroky:
- väzba toxínu na jeho receptor na povrchu bunky;
- zhlukovanie nabitých receptorov do potiahnutých jamiek a internalizácia toxínu prostredníctvom receptorom sprostredkovanej endocytózy; po okyslení endocytového vezikula membránovo asociovanou protónovou pumpou riadenou ATP,
- vloženie transmembránovej domény do membrány a uľahčenie dodania katalytickej domény do cytozolu a
- ADP-ribozylácia EF-2, čo vedie k ireverzibilnému potlačeniu syntézy bielkovín.
Ukázalo sa, že jediná molekula katalytickej domény dodaná do cytozolu je dostatočná na to, aby bola pre bunku smrteľná.
Schematický diagram intoxikácie záškrtom citlivej eukaryotickej bunky.
Toxín sa viaže na receptor na povrchu bunky a internalizuje sa prostredníctvom endocytózy sprostredkovanej receptorom; po okyslení endozómu sa transmembránová doména vloží do membrány vezikula; katalytická doména sa doručí do cytozolu, čo vedie k inhibícii syntézy proteínov a bunkovej smrti.
Epidemiológia
Pred masovou imunizáciou populácie USA difterickým toxoidom bola diftéria zvyčajne detskou chorobou. Pozoruhodným aspektom masovej imunizácie difterickým toxoidom je, že s rastúcim percentom populácie s ochrannými hladinami antitoxínovej imunity (≥ 0,01 IU/ml) klesá frekvencia izolácie toxigénnych kmeňov z populácie. Dnes je v Spojených štátoch, kde klinická diftéria takmer úplne vymizla, izolácia toxigénnych kmeňov C. diphtheriae zriedkavá. Keďže subklinická infekcia už neposkytuje zdroj expozície difterickému antigénu a pokiaľ nie je posilnená, imunita voči antitoxínu klesá, veľké percento dospelých (30 až 60 %) má hladiny antitoxínu pod ochrannými hladinami a je ohrozené. V Spojených štátoch, Európe a východnej Európe sa nedávne ohniská diftérie vyskytli predovšetkým u jednotlivcov, ktorí zneužívajú alkohol a/alebo drogy. V rámci tejto skupiny majú nositelia toxigénnej C. diphtheriae stredne vysokú úroveň antitoxickej imunity. Nedávne zrútenie opatrení verejného zdravotníctva v Rusku viedlo k tomu, že sa záškrt stal epidémiou. Do konca roka 1994 bolo v Rusku hlásených viac ako 80 000 prípadov a viac ako 2 000 úmrtí.
Ohniská záškrtu sú takmer vždy spojené s nositeľom imunity, ktorý sa vrátil z oblasti, kde je záškrt endemický. Nedávne ohniská klinického záškrtu v Spojených štátoch a Európe boli skutočne spojené s cestovateľmi vracajúcimi sa z Ruska a východnej Európy. Toxigénne kmene C. diphtheriae sa prenášajú priamo z človeka na človeka kvapôčkami vo vzduchu. Je známe, že toxigénne kmene môžu priamo kolonizovať nosohltanovú dutinu. Okrem toho sa toxigénny gén môže šíriť nepriamo uvoľnením toxigénneho korynebakteriofága a lyzogénnou transformáciou netoxigénneho autochtónneho C. diphtheriae in situ. [ 5 ]
Okrem stanovenia biotypu a lyzotypu izolátov C. diphtheriae sa teraz na štúdium prepuknutia záškrtu môžu použiť techniky molekulárnej biológie. Na štúdium klinických prepuknutí sa použili vzorce štiepenia chromozomálnej DNA C. diphtheriae reštrikčnými endonukleázami, ako aj použitie klonovaných inzerčných sekvencií korynebakterií ako genetickej sondy.
Schickov test sa už mnoho rokov používa na posúdenie imunity voči difterickému toxínu, hoci v mnohých oblastiach ho v súčasnosti nahradil sérologický test na špecifické protilátky proti difterickému toxínu. Pri Schickovom teste sa malé množstvo difterického toxínu (približne 0,8 ng v 0,2 ml) vstrekne intradermálne do predlaktia (miesto testovania) a 0,0124 μg difterického toxínu v 0,2 ml sa vstrekne intradermálne do kontrolného miesta. Merania sa vykonávajú po 48 a 96 hodinách. Nešpecifické kožné reakcie zvyčajne vrcholia po 48 hodinách. Po 96 hodinách erytematózna reakcia s možnou nekrózou v mieste testovania naznačuje nedostatočnú antitoxickú imunitu na neutralizáciu toxínu (≤ 0,03 IU/ml). Zápal v testovacej aj kontrolnej oblasti po 48 hodinách naznačuje hypersenzitívnu reakciu na antigénový prípravok.
Formuláre
Corynebacterium diphtheriae infikuje nosohltan alebo kožu. Toxigénne kmene produkujú silný exotoxín, ktorý môže spôsobiť záškrt. Medzi príznaky záškrtu patrí faryngitída, horúčka a opuch krku alebo oblasti okolo kožnej lézie. Záškrtové lézie sú pokryté pseudomembránou. Toxín sa šíri krvným obehom do vzdialených orgánov a môže spôsobiť paralýzu a kongestívne zlyhanie srdca. [ 6 ]
Existujú dva typy klinickej diftérie: nazofaryngeálna a kožná. Príznaky faryngeálnej diftérie siahajú od miernej faryngitídy až po hypoxiu v dôsledku obštrukcie dýchacích ciest pseudomembránou. Postihnutie krčných lymfatických uzlín môže spôsobiť silný opuch krku (záškrt typu býčieho krku) a u pacienta sa môže vyvinúť horúčka (≥ 38 °C). Kožné lézie pri kožnej diftérii sú zvyčajne pokryté sivohnedou pseudomembránou. V dôsledku pôsobenia difterického toxínu na periférne motorické neuróny a myokard sa môžu vyvinúť život ohrozujúce systémové komplikácie, najmä strata motorických funkcií (napr. ťažkosti s prehĺtaním) a kongestívne zlyhanie srdca.
Ovládanie
Kontrola záškrtu závisí od adekvátnej imunizácie záškrtovým toxoidom: záškrtový toxín inaktivovaný formaldehydom, ktorý zostáva antigénne neporušený. Toxoid sa pripravuje inkubáciou záškrtového toxínu s formaldehydom pri teplote 37 °C v alkalických podmienkach. Imunizácia proti záškrtu by sa mala začať v druhom mesiaci života sériou troch základných dávok v 4- až 8-týždňových intervaloch, po ktorých nasleduje štvrtá dávka približne 1 rok po poslednom základnom očkovaní. Záškrtový toxoid sa široko používa ako zložka vakcíny proti záškrtu, čiernemu kašľu a tetanu (DPT). Epidemiologické štúdie ukázali, že imunizácia proti záškrtu je účinná približne na 97 %. Hoci sa v Spojených štátoch a Európe praktizuje hromadná imunizácia proti záškrtu a miera očkovania detí je primeraná, veľká časť dospelej populácie môže mať titre protilátok pod ochrannými hladinami. Dospelí by mali byť preočkovaní záškrtovým toxoidom každých 10 rokov. Cestujúci do oblastí s vysokým výskytom endemického záškrtu (Stredná a Južná Amerika, Afrika, Ázia, Rusko a východná Európa) by mali dostať posilňovaciu dávku toxoidom záškrtu a tetanu. V posledných rokoch minimalizovalo používanie vysoko purifikovaných toxoidových prípravkov na imunizáciu občasné závažné reakcie z precitlivenosti. Posilňovacia dávka toxoidom záškrtu a tetanu by mala byť podaná cestujúcim do oblastí s vysokým výskytom endemického záškrtu (Stredná a Južná Amerika, Afrika, Ázia, Rusko a východná Európa). V posledných rokoch minimalizovalo používanie vysoko purifikovaných toxoidových prípravkov na imunizáciu občasné závažné reakcie z precitlivenosti. Posilňovacia dávka toxoidom záškrtu a tetanu by mala byť podaná cestujúcim do oblastí s vysokým výskytom endemického záškrtu (Stredná a Južná Amerika, Afrika, Ázia, Rusko a východná Európa). V posledných rokoch minimalizovalo používanie vysoko purifikovaných toxoidových prípravkov na imunizáciu občasné závažné reakcie z precitlivenosti.
Hoci sa antibiotiká (ako penicilín a erytromycín) používajú ako súčasť liečby pacientov so záškrtom, rýchla pasívna imunizácia difterickým antitoxínom je najúčinnejšia pri znižovaní úmrtnosti. Dlhý polčas rozpadu špecifického antitoxínu v krvnom obehu je dôležitým faktorom pri zabezpečovaní účinnej neutralizácie difterického toxínu; aby však bol antitoxín účinný, musí s toxínom reagovať predtým, ako môže vstúpiť do bunky.
Rekonštrukcia difterického toxínu pre vývoj cytotoxínov špecifických pre eukaryotické receptory
Proteínové inžinierstvo je nová a rýchlo sa rozvíjajúca oblasť molekulárnej biológie; kombinuje metodiky rekombinantnej DNA a syntézu DNA na pevnej fáze s cieľom navrhnúť a skonštruovať chimérické gény, ktorých produkty majú jedinečné vlastnosti. Štúdie vzťahov medzi štruktúrou a funkciou difterického toxínu jasne ukázali, že tento toxín je trojdoménový proteín: katalytická, transmembránová a receptorová. Bolo možné geneticky nahradiť natívnu doménu difterického toxínu viažucu receptor rôznymi polypeptidovými hormónmi a cytokínmi (napr. α-melanocyty stimulujúci hormón [α-MSH], interleukín (IL) 2, IL-4, IL-6, IL-7, epidermálny rastový faktor). Výsledné chimérické proteíny alebo fúzne toxíny kombinujú špecifickosť cytokínu viažuceho receptor s transmembránovými a katalytickými doménami toxínu. V každom prípade sa ukázalo, že fúzne toxíny selektívne otravujú iba tie bunky, ktoré nesú zodpovedajúci cieľový receptor. Prvý z týchto geneticky modifikovaných fúznych toxínov, DAB 389IL-2, sa v súčasnosti hodnotí v klinických štúdiách na ľuďoch na liečbu refraktérnych lymfómov a autoimunitných ochorení, pri ktorých bunky s vysokoafinitnými receptormi IL-2 hrajú dôležitú úlohu v patogenéze.[ 7 ] Podávanie DAB 389 IL-2 sa ukázalo ako bezpečné, dobre tolerované a schopné vyvolať trvalú remisiu ochorenia bez závažných vedľajších účinkov. Je pravdepodobné, že fúzne toxíny na báze difterického toxínu sa stanú dôležitými novými biologickými činidlami na liečbu špecifických nádorov alebo ochorení, pri ktorých je možné zacieliť na špecifické receptory na povrchu buniek.